| |
|
 |
FISIOLOGÍA
Y FUNCIONAMIENTO ÓPTICO DEL OJO |
 |
|
|
|
|
| |
|
|
| |
El
modelo de visión de Kepler interpretó de forma
satisfactoria muchos aspectos del proceso físico de la visión.
Sin embargo, también adolece de limitaciones y
errores propiciados por el desconocimiento en su época de
aspectos sobre la fisiología del ojo y del papel que juega el
cerebro en la construcción de las imágenes. |
|
| |
|
|
| |
 |
|
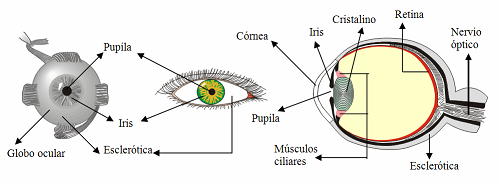
El
diagrama adjunto muestra un esquema del ojo humano que viene a
ser un cuerpo esférico de unos 2’5cm de diámetro.
El orificio por donde entra la luz se llama pupila y tiene un diámetro entre
2mm y
8mm que se regula según la intensidad de luz. La capa más
externa del ojo se llama esclerótica y consta de una
membrana blanca, llamada córnea, que en su zona anterior es abombada y
transparente .
La
primera matización que hemos de hacer al modelo de
visión de Kepler es que, aunque ambos contribuyen a
ello, es la córnea y no el cristalino la que produce casi toda la convergencia de
los haces de luz incidentes. |
|
|
| |
|
|
| |
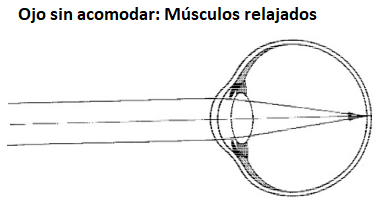
La luz penetra
en el ojo a través de la córnea, atraviesa la pupila y después
el cristalino. Éste está constituido por una materia gelatinosa
(de índice de refracción 1’4) y se puede considerar una lente
biconvexa, que provoca una segunda convergencia. Esta segunda
convergencia realiza una especie de “ajuste fino” del haz, de
tal forma que, mediante un proceso instintivo, llamado
acomodación, se modifica la curvatura del cristalino propiciando
que la imagen se produzca en la retina. En la
acomodación se contrae un
músculo, llamado ciliar, haciendo que disminuya la tensión sobre los
unos ligamentos suspensores que sostienen el cristalino.
|
|
| |
|
|
| |
|
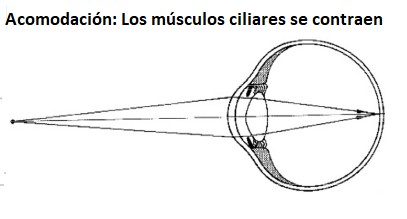
A medida que
disminuye esta tensión, el cristalino se hace más
esférico y así aumenta su potencia refractiva para poder
ver con nitidez objetos próximos. La capacidad de
acomodación del ojo humano está limitada por las
características físicas del cristalino y la capacidad de
contracción del músculo ciliar. Un ojo medio que
funcione bien (ojo emétrope) se puede acomodar para
observar objetos situados entre "el infinito" (el
músculo ciliar está relajado y el ojo presenta la mínima
convergencia) y a 25cm
del ojo (el músculo ciliar relajado propicia la máxima
convergencia). Estos puntos reciben el nombre de
punto remoto y
punto próximo respectivamente. |
|
 |
|
 |
|
|
| |
|
|
| |
|
 |
|
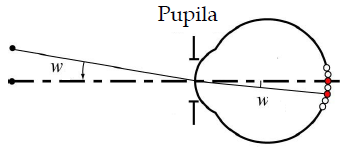
Sea al
visionar objetos próximos u objetos alejados, se llama
agudeza visual a la capacidad del ojo para
separar los detalles más finos de los objetos. Un ojo
estándar u ojo emétrope es capaz de separar las imágenes
de dos puntos que subtienden desde el centro de la
pupila de entrada un ángulo superior a w=1.3´. |
|
|
| |
|
|
| |
|
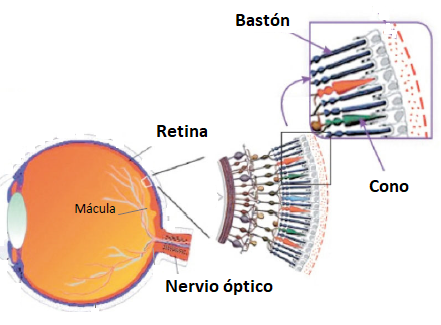
Para la
recepción de la imagen hay en la retina unas células
especializadas, llamadas conos y bastones, que son sensibles a
los rayos luminosos. Los conos, en un número del orden de siete
millones tienen una alta resolución, se activan en buenas
condiciones de iluminación o luz "diurna", y son sensibles a detalles finos de contraste,
al color (mediante visión tricromática R, G, B) y a la forma. La
mayor parte de ellos se encuentra en una depresión cerca
del polo posterior del globo ocular, llamada fóvea. Por su parte, los bastones, en número del orden de
120 millones, tienen baja resolución, se localizan en las partes más periféricas de la
retina, y se activan con iluminación escasa, cuando reciben luz tenue o "nocturna". Los
bastones no son capaces de distinguir el detalle fino o el
color. Por eso, con baja iluminación es difícil distinguir los colores o ver límites
precisos. Al
recibir la luz, ambos
fotorreceptores (conos y
bastones) generan impulsos eléctricos. En la retina existen
otros dos grupos de células que se encargan de
transmitir esos impulsos nerviosos al
cerebro. Los axones de uno de estos grupos forman las fibras del
nervio óptico, que abandona la región posterior del globo ocular
un poco hacia el lado nasal del centro del mismo. Así pues, los
impulsos se propagan al cerebro a través del nervio óptico y es
ahí, en el cerebro, donde se realiza la construcción de la imagen y
se interpreta lo que vemos mediante un proceso bastante complejo. Como
veremos más adelante, nada tiene que ver este proceso con la simple
recepción de una supuesta imagen acabada. |
|
 |
|
|
| |
|
|
| |
| |
Índice |
 |
|
|
|
| |
|
|
|